Page 9 of 29
OG3.1 | Ovulation, Menstruation, Fertilization and Implantation — SDL Guide (Part 2)
Gametogenesis: Oogenesis and Spermatogenesis
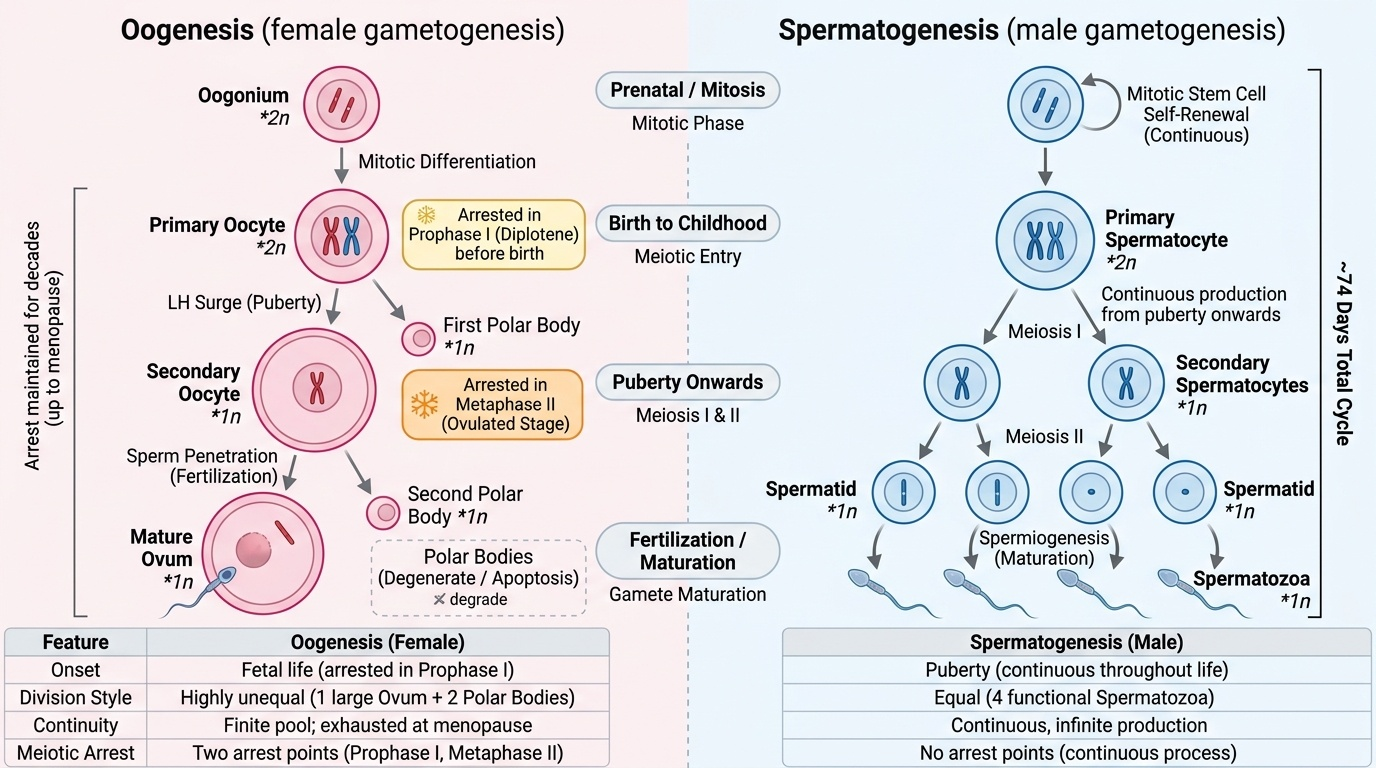
Gametogenesis is the production of haploid gametes (ovum and sperm) from diploid precursors through meiosis. Oogenesis and spermatogenesis share the same meiotic machinery but differ profoundly in timing, location, number of functional products, and regulatory signals — and these differences have direct clinical relevance.
Oogenesis begins during fetal life: oogonia (diploid precursors) in the fetal ovary undergo mitotic proliferation and then enter meiosis I, arresting in the diplotene stage of prophase I before birth. This arrest is maintained throughout childhood and reproductive life, with individual follicles resuming meiosis I only upon exposure to the LH surge — sometimes decades after the arrest began. Meiosis I produces an unequal division: a large secondary oocyte retaining most cytoplasm, and a small first polar body (which may or may not complete meiosis II). The secondary oocyte immediately enters meiosis II but arrests again at metaphase II — this is the stage at which ovulation occurs. Meiosis II completes only when a sperm penetrates the oocyte, producing the definitive mature ovum and the second polar body. The result: each primary oocyte produces only one functional ovum (and two polar bodies, which degenerate).
Spermatogenesis begins at puberty in the seminiferous tubules of the testes and is continuous throughout adult life. Spermatogonia (diploid stem cells) undergo mitotic self-renewal and differentiation into primary spermatocytes, which complete meiosis I to form secondary spermatocytes, and then meiosis II to form spermatids. Spermatids undergo a further maturation process (spermiogenesis) — losing cytoplasm, forming the acrosomal cap and flagellum — to become mature spermatozoa. Each primary spermatocyte produces four functional spermatozoa (equal division of cytoplasm; no polar bodies). The entire process from spermatogonium to spermatozoon takes approximately 74 days.
| Feature | Oogenesis | Spermatogenesis |
|---|---|---|
| Onset | Fetal life (before birth) | Puberty |
| Location | Ovary (follicles) | Seminiferous tubules |
| Duration | Decades (arrested meiosis) | ~74 days |
| Arrest point | Prophase I (at birth); Metaphase II (at ovulation) | No arrest |
| Cytoplasm division | Unequal — one large ovum | Equal — four spermatids |
| Functional products per precursor | 1 ovum + 2 polar bodies | 4 spermatozoa |
| Continuity | Finite pool; exhausted at menopause | Continuous throughout adult life |
Provided image
SELF-CHECK
At which meiotic stage is the oocyte arrested at the time of ovulation, and what event triggers completion of this arrested division?
A. Diplotene of prophase I; triggered by the FSH surge
B. Metaphase I; triggered by corpus luteum progesterone
C. Metaphase II; triggered by sperm penetration of the zona pellucida
D. Anaphase II; triggered by hCG from the trophoblast
Reveal Answer
Answer: C. Metaphase II; triggered by sperm penetration of the zona pellucida
By the time of ovulation, the secondary oocyte has completed meiosis I (triggered by the LH surge) but is now arrested at metaphase II. This arrest is maintained until sperm penetration of the zona pellucida, which triggers completion of meiosis II — producing the mature ovum and the second polar body. If fertilisation does not occur, the oocyte degenerates without completing meiosis II.
Fertilisation: From Ovulation to Zygote Formation
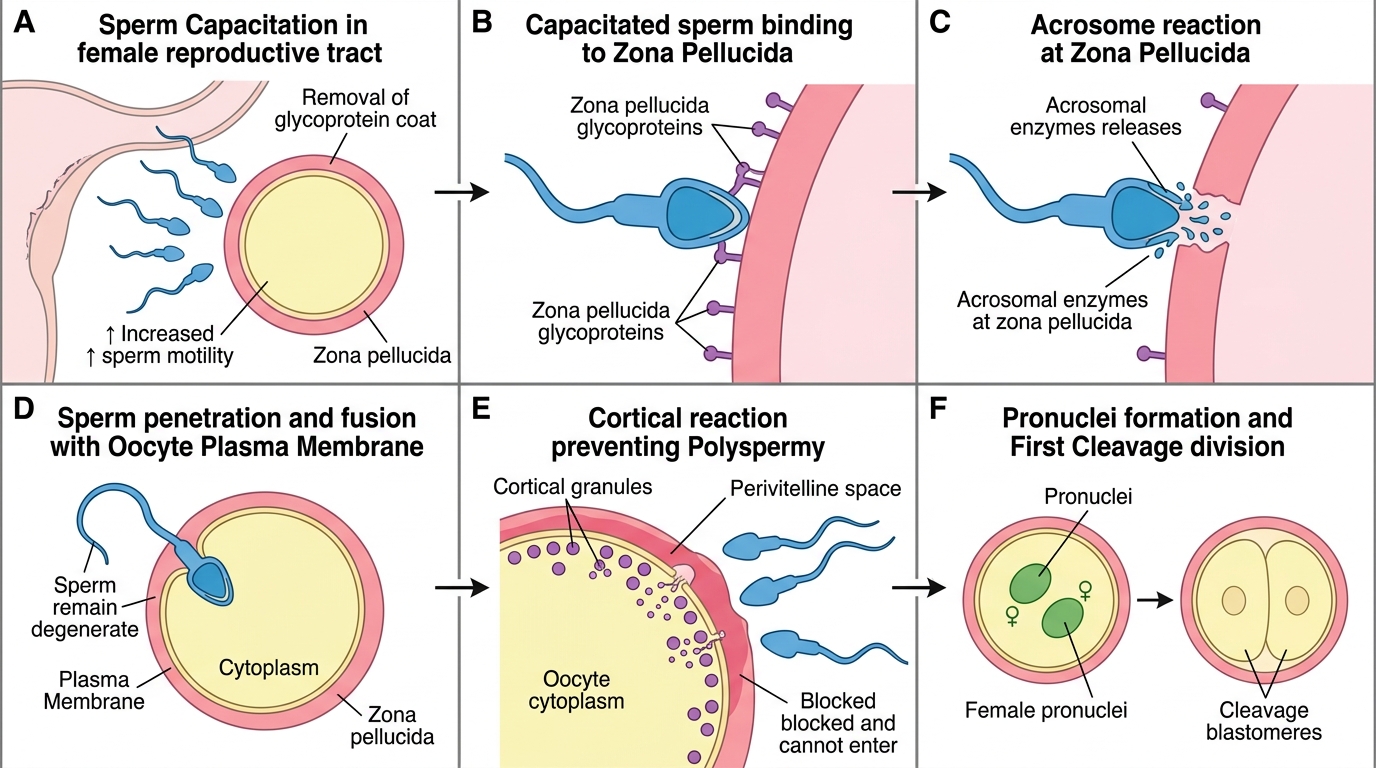
Fertilisation is a sequential, multi-step process that transforms a secondary oocyte and a spermatozoon into a diploid zygote. Each step must succeed; failure at any point results in no conception. The process normally occurs in the ampulla of the fallopian tube within approximately 12–24 hours of ovulation (the window during which the ovum remains viable). Sperm deposited in the vagina can survive in the female genital tract for up to approximately 72 hours, explaining why intercourse up to 3 days before ovulation can result in fertilisation.
Capacitation: Spermatozoa deposited in the vagina are not yet capable of fertilising an oocyte. They undergo capacitation during their transit through the cervical mucus and uterine cavity and fallopian tube — a process involving removal of cholesterol from the sperm plasma membrane, increased flagellar beat frequency, and the ability to undergo the acrosome reaction. Capacitation takes approximately 6–8 hours in vivo.
The acrosome reaction: When the capacitated sperm reaches the cumulus oophorus surrounding the oocyte, its flagellum drives it through the cumulus cells. Upon contact with the zona pellucida (the glycoprotein coat surrounding the oocyte), binding to the ZP3 receptor triggers the acrosome reaction — fusion of the outer acrosomal membrane with the sperm plasma membrane, releasing hydrolytic enzymes (including acrosin) that digest a path through the zona pellucida.
Sperm penetration and the cortical reaction: Once a single sperm penetrates the zona pellucida and fuses with the oocyte plasma membrane, the cortical reaction is immediately triggered — cortical granules beneath the oocyte membrane exocytose, releasing enzymes that modify the zona pellucida (zona hardening or 'zona reaction'), rendering it impenetrable to further sperm. This is the primary block to polyspermy (fertilisation by more than one sperm, which would result in a triploid embryo incompatible with development).
Completion of meiosis II and pronuclei formation: Sperm penetration triggers completion of meiosis II in the oocyte, producing the mature ovum and the second polar body. The sperm head decondenses to form the male pronucleus; the oocyte nucleus forms the female pronucleus. The two pronuclei move together (syngamy) and their chromosomes combine at the first cleavage division — producing the zygote with the normal diploid complement (46 chromosomes, or 23 pairs, one set from each parent).
Step-by-Step Mechanism of Fertilisation
SELF-CHECK
What is the cortical reaction, and what is its primary purpose during fertilisation?
A. The release of acrosomal enzymes by the sperm to penetrate the zona pellucida
B. The exocytosis of cortical granules from the oocyte that hardens the zona pellucida and prevents additional sperm from penetrating
C. The fusion of the male and female pronuclei to restore the diploid chromosome number
D. The depolarisation of the oocyte plasma membrane that triggers completion of meiosis II
Reveal Answer
Answer: B. The exocytosis of cortical granules from the oocyte that hardens the zona pellucida and prevents additional sperm from penetrating
The cortical reaction is the release of cortical granules from beneath the oocyte plasma membrane immediately after the first sperm fuses with the oocyte. The granule contents modify the zona pellucida — making it impenetrable to further sperm — thus preventing polyspermy. This is essential because fertilisation by more than one sperm produces a triploid embryo (69 chromosomes) that cannot develop normally.
Implantation: Endometrial Receptivity and Trophoblast Invasion
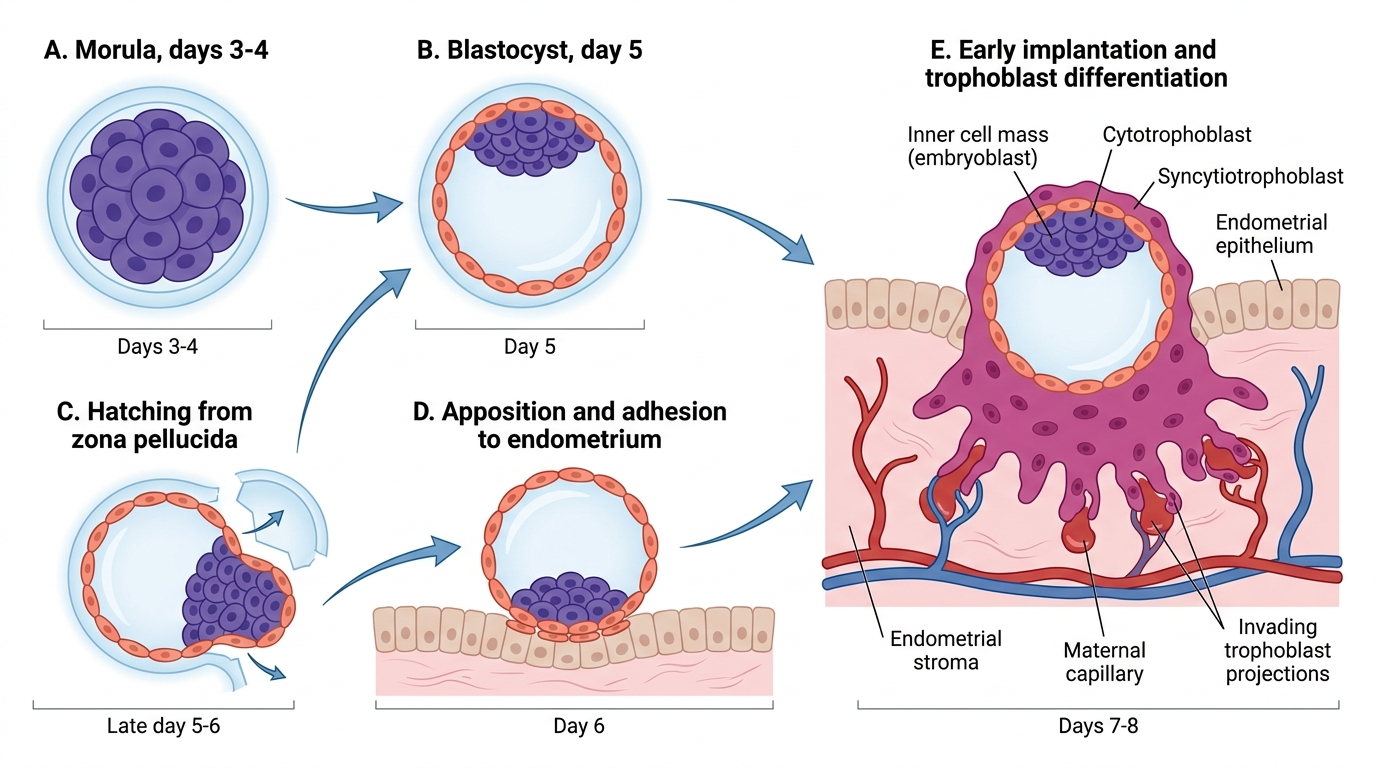
After fertilisation in the ampulla, the zygote begins a series of cleavage divisions as it is transported along the fallopian tube towards the uterus — a journey taking approximately 3–4 days. The cleavage divisions increase cell number without increasing total cell mass, producing progressively smaller cells called blastomeres. By day 3–4 there is a solid ball of 16+ cells — the morula. As the morula enters the uterine cavity, fluid begins to accumulate within it, forming a fluid-filled cavity (blastocoel); this marks the blastocyst stage (approximately day 5). The entire transition from a single-celled zygote to a blastocyst ready for implantation represents a remarkable period of self-directed development, occurring while the embryo is still enclosed within the zona pellucida and floating free in the uterine cavity, entirely dependent on its own metabolic resources and on uterine secretions for nutrition. The timing of this journey is precisely synchronised with endometrial preparation: the endometrium must be in the secretory phase and the implantation window must be open when the hatched blastocyst arrives — a failure of synchrony at this step is a major cause of implantation failure even when a morphologically normal blastocyst is transferred in IVF. The blastocyst has two distinct cell populations:
- Inner cell mass (embryoblast): a cluster of cells at one pole that will become the embryo proper and the yolk sac
- Trophoblast: the outer layer of cells that will form the placenta and extraembryonic membranes
Before implantation can occur, the blastocyst must escape from the zona pellucida — a process called hatching (approximately day 5–6). The blastocyst then 'floats' freely in the uterine cavity for 1–2 days, sampling the endometrial secretions.
Implantation window and endometrial receptivity: Implantation normally occurs approximately 6–10 days after fertilisation (cycle days 20–24 in a regular 28-day cycle), during the narrow implantation window. At this time the endometrium, under progesterone from the corpus luteum, expresses specific adhesion molecules (e.g. integrins, L-selectin ligands) and pinopodes (surface protrusions) that allow trophoblast attachment. Outside this window the endometrium is refractory.
Trophoblast invasion: The blastocyst adheres to the posterior wall of the uterine fundus (most commonly) with its inner cell mass pole facing the endometrium. The trophoblast cells at this pole differentiate into:
- Cytotrophoblast (inner, proliferating, mitotically active layer)
- Syncytiotrophoblast (outer, invasive, multinucleated layer that invades the endometrial stroma and maternal blood vessels)
The syncytiotrophoblast secretes human chorionic gonadotrophin (hCG) from the moment it forms. hCG binds to LH receptors on the corpus luteum, preventing its regression and sustaining progesterone production — maintaining the endometrium and preventing menstruation. Rising hCG is detectable in maternal blood by approximately day 8–9 post-fertilisation (approximately 23–24 days of the menstrual cycle), and in urine by day 10–14 — the basis of the home pregnancy test.
Decidualisation: The endometrial stromal cells, under progesterone, transform into large, polygonal decidual cells rich in glycogen and lipids. This process (decidualisation) produces the decidua basalis (at the implantation site, forming the maternal component of the placenta), decidua capsularis (over the embryo), and decidua parietalis (lining the rest of the uterine cavity). The decidua regulates trophoblast invasion depth and provides nutrition to the early embryo.
Blastocyst Development and Early Implantation